|
|

|
Caminhos da CardiologiaOs seis coraçőes do homem: Um ensaio
Mario Rigatto* Raramente os cientistas relatam os seus estudos obedecendo ŕ cronologia com que os fatos ocorreram e ŕ seqüęncia com que as idéias brotaram em suas mentes. As distorçőes săo compreensíveis em funçăo dos objetivos buscados: arrumar a exposiçăo de forma mais lógica - quem sabe mais bonita - e facilitar o entendimento do leitor. Neste relato procurarei expor as experięncias e as idéias que me levaram ŕ concepçăo de que o homem. possui seis coraçőes, tal como estas experięncias e estas idéias se sucederam no tempo, ao longo destes últimos anos 1.
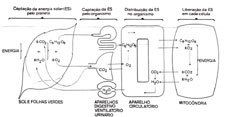
A principal funçăo da circulaçăo é a respiraçăo Há tempo venho me convencendo de que a grande funçăo do aparelho circulatório é assegurar a respiraçăo celular 2-4. A idéia de que o aparelho circulatório serve, eminentemente, a uma funçăo respiratória năo é do agrado dos cardiologistas. Segundo tenho verificado, para eles, a funçăo maior do aparelho circulatório é fazer o sangue circular. O que, em princípio, está certo. Mas dentro da hierarquia das coisas que circulam com o sangue, năo me sobram dúvidas de que săo as exigęncias do transporte das matérias primas da respiraçăo que determinam a bioengenharia do aparelho circulatório. A proposiçăo de que o aparelho circulatório serve mesmo é para respirar, aumenta o meu débito para com os cardiologistas. Débito que se iniciou, há alguns anos, com uma outra proposiçăo que lhes submeti ŕ apreciaçăo: a de que a sede anatômica do amor năo é o coraçăo mas, muito mais provavelmente, o pulmăo 5. Esta proposiçăo inquietou duplamente os especialistas do coraçăo: pelo fato em si e por suas repercussőes econômicas. Uma das justificativas para a consulta dos cardiologistas ser um pouco mais cara do que as dos demais subespecialistas da Medicina Interna, deve-se ao fato de terem eles sob sua responsabilidade, além do centro da circulaçăo, o centro do amor. Os argumentos que apresentei, no entanto, abalaram os meus companheiros de especialidade: "O que pode o amor fazer ao coraçăo?" perguntava-lhes eu. "Acelerar a freqüęncia de seus batimentos?". " Vejam em contraste, a riqueza das manifestaçőes afetivas do pulmăo: é com o pulmăo que rimos, é com o pulmăo que choramos, é com o pulmăo que suspiramos; é do pulmăo que saem todas as interjeçőes afetivas; e todos os ais de amor". E, completava: "O que pode uma discreta taquicardia sinusal, quando comparada ŕ respiraçăo arfante de uma mulher apaixonada?". Năo obstante o desassossego que causa, a proposiçăo de que a circulaçăo serve primariamente para respirar é, também, irretorquível. Se nós pararmos o aparelho circulatório, morreremos em quatro minutos, por falta de oxigęnio. Se por um passe de mágica, com o aparelho circulatório parado, conseguirmos oferecer oxigęnio a todas as células do organismo, morreremos em onze minutos, por excesso de CO, Falta de oxigęnio e excesso de CO2 săo marcas registradas de insuficięncia respiratória. Se num segundo lance de mágica, com a circulaçăo parada, removermos o CO, em excesso, morreremos em tręs semanas, por acúmulo de catabolitos. Uma demonstraçăo eloqüente da primazia dada pelo aparelho circulatório ŕs suas funçőes respiratórias. De acordo com estas idéias, o esquema que tenho proposto para a respiraçăo é o que está representado na figura 1. O fenômeno desenvolve-se em quatro grandes palcos: o meio ambiente, as entradas e as saídas do corpo humano (pulmőes, tubo digestivo e rins), a circulaçăo, e as células, representadas pelas mitocôndrias. No meio ambiente, seis moléculas de água juntam-se a seis moléculas de CO2 para formar uma molécula de glicose. Esta reaçăo, bem conhecida como fotossíntese, só ocorre em presença de energia solar e ao nível de folhas verdes. A importância ímpar da fotossíntese reside no fato de que, ao se formar a molécula de glicose, uma parcela da energia solar é nela envolucrada. E o oxigęnio, simultaneamente produzido, é a "chave" que assegura a oportuna abertura deste envólucro.
Cabe ao aparelho circulatório, na terceira etapa do processo, levar o oxigęnio e a glicose a todas as células do organismo. Ao nível das células, no interior das mitocôndrias, uma molécula de glicose e seis moléculas de oxigęnio reagem entre si. Nesta reaçăo, abre-se a molécula de glicose e dela sai a energia solar de que era portadora. Os subprodutos da reaçăo mitocondrial, seis moléculas de CO, e seis moléculas de água, săo apanhadas pelo sangue e, através, respectivamente, dos pulmőes e dos rins, retornam ao meio ambiente para iniciar uma nova reaçăo de fotossíntese. O processo respiratório, como acima se resume, é o processo escolhido pela Criaçăo para fazer de cada um de nós uma máquina biológica movida a energia solar.
O oxigęnio como modelador do aparelho circulatório Dentro da principal funçăo do aparelho circulatório, transportar as matérias primas da respiraçăo celular, o destaque dado ao oxigęnio é notável. Tenho para mim que a necessidade de pronto, rápido e suficiente transporte de O2, ŕs células seja o fator que modela o aparelho circulatório de cada ser vivo. Sempre me pareceu curiosa a aparentemente paradoxal falta de reserva de oxigęnio no nosso organismo. Quando ficamos privados de alimentos, precisamos de alguns meses para morrer. Quando ficamos privados de água, precisamos de algumas semanas para morrer. No entanto, quando ficamos privados de oxigęnio, morreremos em quatro minutos. Relativamente falando, as reservas de oxigęnio parecem bem menores que as de água e as de alimento. Será verdade? E se for, por quę? Na busca de uma resposta vejo uma maneira de encarar o fenômeno que o torna mais plausível. Se imaginarmos a atmosfera como um grande oceano e a superfície terrestre como o seu fundo, nós seres vivos, estamos mergulhados neste oceano. Ou seja, mergulhados em oxigęnio (fig. 2).
Já a água năo está assim tăo prontamente disponível. Em muitos locais, para obtę-la, é preciso esperar pela chuva; ou encontrar um rio ou lago. O que pode tomar semanas. Os alimentos estăo ainda mais distantes, Algumas vezes, seria preciso esperar que amadurecessem. O que tomaria meses. Uma seqüęncia temporal que se coaduna melhor com a realidade do que a comumente proposta. A fartura de oxigęnio no ambiente em que estamos mergulhados, provę, de modo muito satisfatório a sua pronta disponibilidade para seres vivos de pequenas dimensőes. Como ocorre com os seres unicelulares. Como os protozoários. Neles a achega de oxigęnio é praticamente imediata. Basta cruzar a membrana que os delimita. No entanto, quando as ambiçőes da Criaçăo crescem, e organismos pluricelulares, de maiores dimensőes, se constituem, é necessário dotá-los de um sistema interior que assegure a chegada de oxigęnio ŕs suas muitas células. Células agora năo mais diretamente banhadas pelo oxigęnio. Este sistema interior, para transporte de oxigęnio, é o aparelho circulatório. É fácil imaginar que o aparelho circulatório, concebido primariamente para levar oxigęnio ŕs células, fique subordinado, em sua concepçăo estrutural e funcional, ŕs exigęncias de disponibilidade de O2, regidas pelo binômio distância-consumo. Em outras palavras: Quanto maior o animal a ser oxigenado, maior a distância a ser percorrida para chegar a cada célula e maior a exigęncia sobre a circulaçăo. Por outro lado, quanto maior o consumo de O2 de cada célula, maior também a exigęncia circulatória por ela imposta.
O pulmăo como bomba circulatória Nesta seqüęncia de perplexidades e proposiçőes um outro elemento passou a me influenciar de forma importante: a constataçăo de que o pulmăo năo bombeia apenas ar: bombeia sangue também. E sua atividade como bomba circulatória tem uma expressăo quantitativa importante 6,7. Nas minhas atividades como pesquisador, tenho estado interessado na circulaçăo capilar pulmonar, Um dos projetos que contemplei e desenvolvi foi o aprimoramento de um método ventilatório para medir o débito desta circulaçăo capilar. O que, na prática, equivale a medir a débito cardíaco 8,9. Através de diferentes abordagens, acabei conseguindo uma técnica ventilatória que atendia bem ao meu desejo. Um método que, em última instância, era um aperfeiçoamento do método desenvolvido por Krogh e Lindhard, em 1912 10. Este método depende da absorçăo, pelo sangue que passa pelo pulmăo, de um gás nele solúvel. O óxido nitroso, N2O, é o gás que tem merecido a preferęncia, dos pesquisadores. Sabendo-se a solubilidade do N2O no sangue e medindo-se o volume dele absorvido através dos pulmőes, na unidade de tempo, pode-se calcular a quantidade de sangue que passou pelos pulmőes, naquele tempo. As limitaçőes da técnica de Krogh e Lindhard eram fundamentalmente duas: a primeira, a dificuldade para determinar o momento em que se alcança equilíbrio entre a concentraçăo de N2O no balăo, no qual o paciente respira, e em seus alvéolos. De maneira que a amostragem do balăo passa representar a amostragem dos alvéolos. Só a partir do momento em que este equilíbrio é atingido é que a mensuraçăo do débito pode ser feita sem erro sistemático. A segunda dificuldade, era demarcar o momento em que começava a recirculaçăo do sangue pelos pulmőes. O problema da recirculaçăo é que ela traz consigo parte do óxido nitroso absorvido quando da primeira passagem do sangue pelos pulmőes. Este sangue, ainda contendo algum óxido nitroso, já năo absorve a mesma quantidade de N2O que absorvia da primeira feita. As mensuraçőes, nestas condiçőes, dăo um débito artificialmente baixo. A minha contribuiçăo foi, usando os melhores recursos tecnológicos da atualidade, encontrar maneiras para delimitar com precisăo e simplicidade os dois pontos críticos acima discutidos8,11. Para tanto, oferecíamos ao paciente uma mistura contendo aproximadamente 15% de óxido nitroso (N2O), 45% de nitrogęnio (N2) e 40% de oxigęnio (O2). Utilizávamos um balăo de borracha, do tipo usado pelos anestesiologistas. O paciente era convidado a respirar a mistura gasosa contida neste balăo, usando uma freqüęncia respiratória maior que a normal, num volume de ar corrente também maior que o normal, para permitir um mais rápido equilíbrio entre o ar dos seus pulmőes e o do balăo (fig. 3 e 4). A composiçăo da mistura gasosa, ao nível da boca, era determinada por amostragem contínua, Ievada a um espectrômetro de massa, para análise. Quatro gases eram continuamente medidos o óxido nitroso, o nitrogęnio, o oxigęnio e o vapor d'água.
Com a introduçăo destes dois pontos de reparo, o método de Krogh e Lindhard pode ser aplicado sem erros sistemáticos, inclusive durante exercício, quando, pelo aumento da velocidade circulatória, o segmento temporal adequado para a mensuraçăo do débito é proporcionalmente menor. Os resultados por nós obtidos para o débito cardíaco mostraram que o método, assim aperfeiçoado, levava a resultados comparáveis ao obtido por cateterismo cardíaco aplicando-se o princípio de Fick 8,9 .
Nossa interpretaçăo é de que a bomba ventilatória pode adicionar importante volume ŕ circulaçăo sangüínea sistęmica. Este aumento é normalmente utilizado durante exercício em funçăo do aumento do volume de ar corrente e da freqüęncia ventilatória que, espontaneamente, ocorrem nesta circunstância. Em face de crescentes cargas de trabalho. esta incorporaçăo de fluxo sangüíneo, gerado pela bomba ventilatória, decresce gradativamente até um ponto em que novos aumentos da freqüęncia ventilatória e do volume de ar corrente nada mais acrescentam ao débito circulatório. É interessante observar que este ponto é o que corresponde ao limiar anaeróbio: cerca de 3.000 ml de consumo de oxigęnio por minuto, VO2, em indivíduos fisicamente aptos mas sem treino atlético (fig. 8). A observaçăo de um aumento do débito sangüíneo pulmonar por influęncia de manobra ventilatória, năo é originalmente nossa. Coube-nos, no entanto, o privilégio de poder determinar quantitativamente este aumento e documentá-lo, năo só em repouso mas também durante exercício, o que, até entăo, năo havia sido possível.
O fenômeno, em si, foi proposto, primeiro por Krogh e Lindhard, em 191210. Năo obstante a objeçăo de alguns grandes pesquisadores da época, entre eles Haldane13 e Henderson14, estudos posteriores demonstraram que a proposiçăo de Krogh e Lindhard era correta. Assim, Daly, em 1927 15 demonstrou que, aplicando sucçőes rítmicas a uma preparaçăo coraçăo-pulmăo, era possível dobrar o débito cardíaco. Em 1951 Baxter e Pearce, trabalhando com căes, documentaram, através de mensuraçăo direta do flux o sangüíneo, aumento do volume sistólico direito em cada movimento inspiratório 16. O aumento do fluxo sangüíneo pulmonar, em face do aumento dos movimentos ventilatórios, foi atribuído a vários fatores: demanda metabólica dos músculos ventilatórios, redistribuiçăo do débito sangüíneo em funçăo da queda da PaCO2, reflexos vasculares 6,7. Os nossos estudos, realizados em tempo inferior ao da recirculaçăo sangüínea pulmonar, permitem excluir as duas primeiras pretensas justificativas. A participaçăo de reflexos vasculares năo pode ser eliminada. Năo obstante, a correlaçăo observada entre os movimentos ventilatórios e o fluxo sangüíneo pulmonar, e as variaçőes desta correlaçăo com o decúbito (menor em decúbIto supino, quando o pulmăo está mais engurgitado) e com o exercício (progressivamente menor em face de cargas crescentes de trabalho quando a turgęncia pulmonar é progressivamente maior) fortemente sugerem que o aumento se deva, no todo ou, pelo menos, em sua maior parte, ŕ uma açăo mecânica da bomba ventilatória.
Inflaçăo de coraçőes A documentaçăo da grande importância da bomba ventilatória sobre o débito sangüíneo nos levou a várias consideraçőes. A primeira foi a percepçăo de que se pode comparar a bomba ventilatória a um legítimo "coraçăo". Entendendo-se como "coraçăo" qualquer compartimento vascular com variaçőes rítmicas de suas dimensőes e com válvulas capazes de direcionar o fluxo do sangue nele contido (fig. 9).
Por analogia, pareceu-nos também claro que a cavidade abdominal, com a veia cava inferior, constituía um outro "coraçăo". A veia cava inferior constitui um compartimento vascular que varia ritmicamente suas dimensőes - em funçăo das variaçőes cíclicas da pressăo que o cerca e decorrentes dos movimentos diafragmáticos - e limitado, de um lado, pelas mais altas válvulas das femorais, que năo permitem o retrocesso do sangue, e, de outro lado, pela válvula tricúspide, que lhe dá plena vazăo. Raciocínio análogo deixa também claro que os membros inferiores possuem um perfeito "coraçăo": veias valvuladas, cercadas por poderosa massa muscular que, ao se contrair, propele o sangue nelas contido. Nesta altura dos acontecimentos, com tamanha inflaçăo de coraçőes, senti necessidade de anal;sar o sistema cardiovascular ao longo de sua evoluçăo filogenética, em busca de subsídios para o modelo que estava vislumbrando: um modelo dotado năo apenas de uma única bomba central, mas de múltiplas bombas. centrais e periféricas, dispostas em seqüęncia - uma circulaçăo com muitos "coraçőes". Revisăo filogenética Para atender ao propósito maior da circulaçăo, transportar eficientemente oxigęnio, a Criaçăo concebeu diversos mecanismos circulatórios, de acordo com a complexidade do organismo a ser por eles servido17. A figura 10 ilustra os modelos fundamentais de circulaçăo18. Assim, para seres unicelulares, como os protozoários, correntes protopIásmicas constituem uma circulaçăo suficiente já que a distância a ser percorrida é pequena e o consumo de oxigęnio năo é grande. Para seres pluricelulares, mas ainda de concepçăo singela, como esponjas e celenterados, o movimento da água na qual estes seres vivem, ao longo das anfratuosidades de seus corpos, constitui mecanismo circulatório adequado.
Numa etapa adiante, em seres um pouco mais complexos, esboça-se a necessidade de um aparelho vascular. Nos crustáceos, um aparelho rudimentar, com vasos, como artérias, leva o sangue a seios ou lagos venosos. Nos oligocetas, aparecem capilares e velas que tendem a "fechar" o aparelho circulatório. Com o surgimento do sistema vascular apareceu, simultaneamente, a bomba propulsora do sangue, nele contido. Apareceu o coraçăo. Estes coraçőes, centrais ou sistęmicos, assumem uma polimorfa apresentaçăo anatômica: podem ser simples vasos pulsáteis, como na minhoca. Podem ser coraçőes tubulares, como nos artrópodos. Ou podem ser coraçőes multicavitários, com duas, tręs ou quatro cavidades, como nos vertebrados. Ŕ medida que o binômio distância-consumo de O2 torna-se mais exigente, um crescente aumento da pressăo circulatória faz-se imprescindível, exigindo um progressivo fechamento do sistema vascular. O que já ocorre nos oligocetas, nas sanguessugas. E progride ŕ medida que se sobe na escala animal. Mas nunca chega a ser total. No próprio homem ainda temos vestígios da circulaçăo lacunar: na circulaçăo venosa cerebral, no baço e no fígado. Além do estabelecimento de um sistema vascular e de seu progressivo fechamento, a natureza utilizou outros recursos para manter o aparelho circulatório adequado ŕs crescentes exigęncias da filogenia. Um destes outros recursos foi a inervaçăo do sistema. Assim, até os ciclóstomos, que constituem os peixes mais primitivos, năo há qualquer evidęncia de inervaçăo do sistema circulatório. Mas dos ciclóstomos para cima, já abrangendo os peixes ósseos, o vago se faz presente. A partir dos répteis, o simpático é adicionado, mantendo-se os dois, vago e simpático, daí por diante. Outro recurso importante foi a introduçăo de válvulas no sistema venoso, para permitir o ortostatismo. Que começa nas espécies logo acima dos peixes. A rede linfática é outro recurso mobilizado. Aparece nos anfíbios e nos répteis, quando ainda se confunde com o sistema venoso. De fato, nestas espécies, há freqüentes desaguadouros da rede linfática no sistema venoso. O ducto torácico, como desaguadouro central, só aparece nos vertebrados superiores. Do ponto de vista das minhas indagaçőes, o mais notável dos aperfeiçoamentos introduzidos no sistema circulatório pela Criaçăo, ao longo da escala filogenética, foram os coraçőes acessórios. Seu aparecimento data do momento em que a propulsăo do sangue, particularmente para garantir o seu retorno ao coraçăo principal, passou a se constituir em desafio. Os anelídeos e os insetos possuem vários coraçőes. Nos mixinoides, peixes dos mais primitivos, há uma grande quantidade de coraçőes: além do coraçăo sistęmico, um coraçăo caudal, um coraçăo cardinal, um coraçăo portal e, ainda coraçőes branquiais. Coraçőes acessórios também săo descritos no sistema linfático em répteis e anfíbios. Os coraçőes acessórios possuem as mais varia. das estruturas anatômicas, tendo, apenas, como denominador comum, competęncia para propelir sangue. Assim, nos cefalópodos, os coraçőes acessórios săo constituídos por tecido músculoepitelial esponjoso que envolve os pequenos vasos. Nos mixinoides, por músculo estriado ao nível das guelras ou por um engenhoso coraçăo músculo-cartilaginoso na cauda (fig. 11). Os coraçőes linfáticos de anfíbios e répteis, săo constituídos por alargamentos de vasos linfáticos, providos de inervaçăo espinal. O corte desta inervaçăo determina a paralisaçăo destes coraçőes, podendo haver, subseqüentemente, movimentos isolados e algo desordenados dos mesmos. Na apreciaçăo global desta evoluçăo filogenética há um ponto que desejo destacar. É o fato de que, uma vez Instalado um novo recurso, o mesmo é mantido nas espécies subseqüentes, mais diferenciadas.
De acordo com a bibliografia atualmente disponível, o único progresso que parece ter sido implantado no sistema circulatório, ao longo de sua evoluçăo filogenética, e depois dele retirado, săo os coraçőes acessórios. Será verdade? Será verdade que um mecanismo tăo sensato, permitindo que bombas estrategicamente localizadas, algumas delas especificamente dedicadas ao atendimento de demandas metabólicas maiores, como o exercício, bombas que tornam a circulaçăo energeticamente mais barata e mecanicamente mais eficiente, haveriam de desaparecer exatamente quando a demanda circulatória atinge o seu maior grau? Ou será, apenas, que os coraçőes acessórios assumiram novas roupagens e, assim, escaparam ŕ nossa percepçăo como entidades anátomo-funcionais individualizadas? Esta última interpretaçăo é a que me parece correta. Creio haver suficiente riqueza e elementos para se reconhecer no nosso sistema circulatório a presença de vários coraçőes acessórios.
Os seis coraçőes do homem De acordo com esta interpretaçăo, acredito que o homem possua seis coraçőes (fig. 12) 1,20. Em seqüęncia, e partindo de uma extremidade, por exemplo, dos
membros inferiores, podemos reconhecer: o "coraçăo periférico% constituído
pelas veias valvuladas dos membros inferiores, cercadas de músculos que se
contraem ritmicamente com os movimentos destes membros; o O "coraçăo periférico" seria o único a atuar exclusivamente durante exercício físico. Os demais atuariam sempre.
Encanto e dúvidas O modelo proposto me encantou. Mas suscitoume também muitas dúvidas. Assim, o "coraçăo periférico" , descrito para os membros inferiores, seria igualmente válido para os membros superiores, embora a valvulaçăo das veias dos membros superiores seja bem menos perfeita que a dos inferiores. A analogia entre os dois, no entanto, é marcante. Por isso, chamo de "coraçăo periférico" tanto o dos membros inferiores quanto o dos membros superiores. Năo me parece razoável dar nomes distintos, a cada um já que as suas características săo fundamentalmente as mesmas: é óbvio que estes "coraçőes periféricos" podem atuar isolada ou conjuntamente, dependendo do número de membros que esteja executando exercício. É importante salientar que apenas o exercício năo-isométrico é capaz de acionar estes coraçőes. Tive dúvidas se o "coraçăo pulmonar" e o "coraçăo abdominal" deveriam ser considerados como dois coraçőes, uma vez que o elemento motor principal é comum a ambos: o diafragma. Parece-me, no entanto, que a riqueza de características próprias a cada um destes "coraçőes" justifica a sua individualizaçăo em separado. Tive dúvidas se as aurículas năo constituem coraçőes em separado. A fibrilaçăo auricular, que anula sua contribuiçăo hemodinâmica, reduz em 10 a 20% o débito sangüíneo. Pareceu-me, no entanto, mais apropriado considerá-las como estruturas Potencializadoras dos coraçőes ventriculares. O último dos coraçőes que aceitei, e que me deixou por mais
tempo reticente, foi o "coraçăo arterial". Acabei, no entanto,
concluindo pela sua validade. A evidęncia em seu favor começa com os peixes teleósteos. Em face da necessidade de potencia. lizar o
coraçăo sistémico, tendo pela frente duas redes capilares a serem vencidas, a
rede capilar branquial e a rede capilar sistęmica, a natureza equipou o
coraçăo destes peixes com um "bulbus arteriosus" (fig. 13). Que é,
legitimamente, um
Ilaçőes fisiopatológicas O modelo proposto para o aparelho circulatório, constituído por uma seqüęncia de bombas aspirante. prementes, possui aspectos interessantes, quando analisado ŕ luz da fisiopatologia circulatória e respiratória. Assim, faz excelente sentido a idéia de que, ao passar do repouso para o exercício, quando se passa a exigir um alto débito sangüíneo para os membros, estes próprios membros, passem a gerar a circulaçăo por eles exigida. Estudos da equipe do Prof. Guyton21 mostram que, em face de estímulos elétricos que aceleram a contraçăo rítmica dos músculos dos membros de animais de laboratório, o retorno venoso, por estes membros, aumenta proporcionalmente. Estas experięncias documentam, objetivamente, o que se propőe como comportamento dos membros do ser humano. É interessante considerar que o movimento do pé exige flexőes mais acentuadas quando caminhamos num aclive do que num declive. No aclive precisamos vencer uma carga maior; conseqüentemente, precisamos de um débito maior. No declive, o oposto sucede. Talvez a grande dificuldade que encontram as mulheres para caminhar depressa e, ainda mais, para correr, quando com saltos altos, dependa mais da relativa imobilizaçăo da bomba circulatória periférica, que os movimentos dos pés representam, do que da incômoda, ainda que elegante, postura musculo-esquelética que os saltos determinam. Qualquer um de nós sabe que, ao correr, basta um aumento de meio centímetro na altura do salto do tęnis para que nossa eficięncia diminua. Pergunto-me se as limitaçőes para a marcha das pessoas com pé chato dependem somente de problemas ortopédicos; ou, se năo também, da menor eficięncia da sístole podálica. O "coraçăo abdominal" possui aspectos interessantes nestas correlaçőes fisiopatológicas. É sabido dos cirurgiőes e dos clínicos que as operaçőes de abdômen alto isto é, operaçőes que determinam incisőes próximas ao diafragma, prejudicam a eficięncia respiratória dos pacientes tanto quanto as operaçőes do tórax. A explicaçăo năo é difícil se nos dermos conta de que estas operaçőes neutralizam, pela dor da incisăo, o "coraçăo abdominal" com a mesma eficięncia com que neutralizam o "coraçăo pulmonar". É um fato curioso, na clínica pneumológica, a extrema dispnéia dos enfisematosos em face do exercício físico quando os bronquíticos, em média, pacientes bem mais graves, toleram exercício equivalente. Será que a retificaçăo das cúpulas diafragmáticas, própria dos enfisematosos. desligando o "coraçăo abdominal" e o "coraçăo pulmonar", năo está na raiz do fenômeno? Um bronquítico mesmo em quadro avançados. mantém boa curvatura das suas cúpulas diafragmáticas, a menos que possua associadamente, um importante componente enfisematoso. É também digna de inquisicăo a possibilidade dos movimentos cíclicos da pressăo intraperitonial influenciarem o retorno venoso hepático. Ŕ medida que aprendemos mais sobre a fisiologia do exercíco. sabemos que um atleta treina o seu fígado tanto quanto treina os seus músculos. O crescimento da circulaçăo hepática parece ser característica do bom condicionamento físico. Será que um bem treinado "coraçăo abdominal", com mais amplas e mais vigorosas excursőes diafragmáticas com uma mais eficiente contraçăo da musculatura da parede anterior do abdômen, năo assegurará melhor retorno venoso ŕ circulaçăo hepática? O "coraçăo arterial" também leva a digressőes. Mellerowicz - citado por Jokl23 - verificou que o aumento da pressăo arterial sistólica, acima dos 50 anos de idade, praticamente năo se observa em homens com atividade física regular. Será que o treinamento físico assegura um bom treinamento ŕ elasticidade das paredes arteriais? Analisados em conjunto, os "coraçőes" conduzem ainda, a outras indagaçőes. É prática corrente, nestes últimos anos, o estabelecimento de programas de treinamento físico para indivíduos que sobrevivem a um infarto do miocárdio 24,25 . Todos os que se ŕ dedicado ŕ observaçăo destes programas de treinamento, impressionam-se com a sensível melhoria circulatória dos pacientes, em face do exercício físico. Năo menos impressionados firam, também, com os discretos progressos que se conseguem documentar no desempenho do coraçăo e que năo guardam proporçăo com o progresso global (26). Será que năo faz melhor sentido imaginar, que um indivíduo, no pós infarto, ao ser colocado em treinamento físico, tem muito mais possibilidades para desenvolver m potencialidades de seus "coraçőes acessórios" do que para melhorar, substancialmente, o seu coraçăo central, já tăo marcado pela doença? Será que um coraçăo empedernido pela fibrose miocárdica e pela aterosclerose coronária, terá mesmo possibilidade de uma grande melhoria em seu desempenho? O mais provável, no meu entender, é que substancial parte do progresso observado se deva, ŕ melhora dos coraçőes periféricos. Que, como regra, nesta nossa civilizaçăo moderna, săo mantidos ociosos e, conseqüentemente, em má forma, porque năo treinados, em funçăo da vida sedentária. A concepçăo aqui proposta facilita também o entendimento das importantes diferenças que existem no comportamento cardíaco de atletas que se dedicam a exercícios isométricos e năo-isométricos. Sabe-se que a prática de exercícios isométricos em pouco ou nada altera o desempenho cardíaco. Assim, nos levantadores de peso, a freqüęncia cardíaca praticamente năo se altera, isto é. năo se reduz com a prática continuada do exercício23. Năo é difícil, no entanto. verificar que o exercício isométrico năo tem possibilidades para um adequado treinamento dos coraçőes periféricos. No campo esportivo, um outro exemplo sobe ao plano das atençőes, em face deste novo modelo circulatório. É conhecimento cada vez mais bem documentado que o bom atleta, do ponto de vista circulatório. É aquele que possui um bom retomo venoso. Analisando-se os desafios mecânicos da circulaçăo, năo é difícil perceber que maior problema é fazer o sangue retomar ao centro do que, do centro, fazę-lo chegar ŕ periferia. Para o transporte do centro ŕ periferia, utilizamos um grande gradiente pressórico. Mas ficamos na obrigaçăo de conseguir o retomo da quantidade levada ŕ periferia com um gradiente pressórico várias vezes menor. Os alemăes documentaram, melhor que quaisquer outros. o fato de que, em face de exercício regularmente praticado, o coraçăo aumenta de peso, aumenta sua massa muscular24,27. Este aumento, como regra năo ultrapassa 50% da massa cardíaca de um indivíduo sedentário. Mas é de substancial importância. O curioso neste aumento é que ele prepondera ro coraçăo direito27. Isto é, no coraçăo que tem diretamente sob sua tutela o retorno venoso. Tudo parece indicar que o adequado treinamento dos coraçőes periféricos tem muito a ver com a excelęncia deste retorno.
Somatório De há muito, e em diferentes estudos, se tem propalado a açăo facilitadora do retorno venoso, criada pela negatividade pleural. O mesmo se tem dito da contratura dos músculos dos membros inferiores. Da importância da pressăo arterial que atinge o leito capilar periférico e da qual deriva a fundamental "vis a tergo". E assim por diante. Năo houve, no entanto, até hoje, a caracterizacăo destes mecanismos nos termos que agora proponho. Acho que a individualizaçăo das várias unidades propulsoras de sangue, como "coraçőes" dispostos em seqüęncia, contribui para uma melhor visăo do conjunto e um melhor entendimento das partes. Seguramente será possível desenvolver métodos que permitam quantificar a contribuiçăo de cada um desses "coraçőes", a exemplo do que já tivemos oportunidade de fazer em relaçăo ao "coraçăo pulmonar". Estas informaçőes talvez nos permitam orientar melhor os programas que prescrevemos visando desenvolver ou recuperar a eficięncia do aparelho circulatório e, através dele, a competęncia respiratória.
Summary The results obtained by the author with a method of his own, capable to determine the influence of the ventilatory movements on the pulmonary blood flow, led him to postulate the existence of a "pulmonary heart". This heart would be represented by the pulmonary vascular compartment limited by the pulmonic valve, its entance, and by the mitral valve, its outlet. Inspiration, distending the compartment , would induce a diastolic phase. Expiration, compressing the compartment, would induce a systolic phase. The measurements made showed a significant increase of the pulmonary blood flow proportional to the intensification of the ventilatory movements. The individualization of a "pulmonary heart" induced the authors to configurate, by analogy, other vascular compartments with blood pumping activity: the "abdominal heart", formed by the inferior vena cava, distended and compressed by the diaphragmatic movements; the "peripheral heart", in the legs or in the arms, formed by the veins of the limbs, compressed by the contraction of the muscles which surround them; and the "arterial heart", formed by the arterial tree through the distension and retrac tion of its elastic walls. All these compartments comply with the basic definition of a "heart": a compartment with cyclic variations of volume, and inbuilt valves to direcblood flow. These heats, added to the two classical "ventricular hearts", set the six sequential hearts model proposed by the author for the human circulatory apparatus. The author analyses the validity of this model in the light of the phylogenetic evolution of the cardiovascular apparatus and of the physiological and pathophysiological illations it enhances.
Referęncias 1. Rigatto, M. - os seis coraçőes do homem (conferęncia). XVI Congr. Bras, de Fisiologia. Resurnos. Rio de Janeiro. 1982. P. 224. 2. Rigatto, M. - FisiopatOlogia da circulagao pulmonar, BykProcienx. Sdo Paulo. 1973, P. 62. 3. Rigatto, M. - Insufici6ncia Pulmonar. In Tarantino, A. B. -Doenqas Pulmonares. Guanabara-Koogan. Rio de Janeiro. 1976. P. 216. 4. Rigatto, M. Insufici6ncia respirat6ria ndo-pulmonar. In Parsioe, C. Conceitos autais em respiragdo. Eyk-Procienx. Săo Paulo. 1978. P. 79. 5. Rigatto, M. - O estudo, o trabalho, nós e o tempo. In Médicos e Sociedade. Byk-Procienx, Săo Paulo. 1976. P. 66. 6. Rigatto. M. - The contribution of the ventilatory pump to venous return during exercise. International Conference on Sports Cardiology: Abstracts Book. Rome. 1978. P, 15. 7. Rigatto. M. - The contribution of the ventilatory pump to venous return. In Lubich, T.; Venerando, A. - Sports cardiology. Aulo Gaggi. Bologna. 1980. P. 75. 8. Rigatto, M. - Mass spectrometry in the study of pulmonary circulation. Bull. Physio-Pathol. Respirat. 3: 473, 1967. 9. Rigatto, M. - Determinaçăo do débito cardíaco pela respiraçăo de óxido nitroso em circuito fechado. Arq. Bras. Cardiol. 21 (supl. 1): 1, 1968. 10. Krogh, A.; Lindhard, J. - Measurements of blood flow thorugh the lungs of man. Skand. Arch. Physiol. 27: 100, 1912. 11. Rigatto, M.; Jones. N. L.; Campbell, E. J. M. - Pulmonary recirculation time: Influence of posture and exercise. Clin. SOL 35: 183, 1968. 12. Rigatto, M.; Turino, G. M.; Fishman, A. P. - Determination of the pulmonary capillary blood flow in man. Circul. Res. 9: 945, 1961. 13. Haldane, J. S. - Respiration, Yale University Press. New Haven. 1922. 14. Henderson, Y. - Volume changes of the heart. PhysioL Rev. 3: 165, 1923. 15. Daly, I. de B. - Negative pressure pulmonary ventilation in the heartlung preparation. J. Physiol. 63: 81, 1927, 16. Baxter, I. G.; Pearce, J. M. - Simultaneous measurements of pulmonary arterial flow and pressure using condenser manometers. J. Physiol. 115: 410, 1951. 17. Johansen, X.; Martin, A. W. - Comparative aspects Of cardiovascular function in vertebrates. In Handbook Of Phy , siology. Hamilton, W. F. - American Physiological Societv. Washington (D.C.) 1965. Vol. III. P. 2583. 18. Prosser, C. L. - Circulation of body fluids. In Comparative animal physiology, S.a ed.. Saunders. Philadelphia. 1973 P. 822. 19. Greene, C. W. - Contributions to the physiology of California hagfish, Polistotrema strouti 1. The anatomv, and physiology of the caudal heart. Am. J. Physiol. 3: 366, Igor), 20. Rigatto, M. - The six hearts of man during exercise. World Scientific Congress "Sport in Modem Society". Abstracts. Book. Thlissi (URSS). 1980. p. 105. 21. Guyton, A. C. - Regulation of venous return. In Cardiac output and its regulation. Saunders. Philadelphia. 1963. p. 163. 22. Johansen, K. - Cardiac output and pulsatile aortic flow in the teleost, Gadus morhua. Comp. Bloch. Physiol. 7: 169. 1962. 23. Jokl, E. - Heart and sport. Thomas. Springfield (Illinois, USA). 1964. 24. Rigatto, M. - Longevidade e atividade fisica. Rev. Ass. Mdd. R. G. Sul 10: 35, 1965. 25. Rigatto, M. - O exercicio na ribalta cardioldgica. Herói ou vilăo? In Médicos e Sociedade. Byk-Procienx. Săo Paulo. 1976. P. 107. 26. IArsen, O. A.; Malmborg, R. O. - Coronary heart disease and physical fitness. Munksgaard. Copenhagen. 1971, 27. Lubich, T.; Venerando, A. - Sports cardiology. Aulo Gaggt. Bologna. 1980.
* Professor Adjunto de Medicina Interna. LTFRGS. Pesquisador Titular,
|
|||||||||||||||||||||||||||||||||||||||||||||||||||


 No pulmăo dá-se a captaçăo do oxigęnio, produzido pela
fotossíntese, e a sua transposiçăo para o sangue. No tubo digestivo dá.se a
captaçăo da glicose, também produzida pela fotossíntese, e a sua
transferęncia, também para o sangue.
No pulmăo dá-se a captaçăo do oxigęnio, produzido pela
fotossíntese, e a sua transposiçăo para o sangue. No tubo digestivo dá.se a
captaçăo da glicose, também produzida pela fotossíntese, e a sua
transferęncia, também para o sangue. Visto deste ângulo, o oxigęnio deixa de ser o menos
disponível dos elementos exigidos para o nosso metabolismo vital. Pelo
contrário, passa a ser o mais disponível. E, em maior quantidade. Dia e noite.
Em qualquer lugar.
Visto deste ângulo, o oxigęnio deixa de ser o menos
disponível dos elementos exigidos para o nosso metabolismo vital. Pelo
contrário, passa a ser o mais disponível. E, em maior quantidade. Dia e noite.
Em qualquer lugar.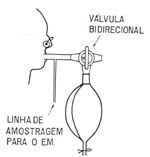 Como se vę na figura 5, o nitrogęnio, que imediatamente antes
do teste se encontrava nos pulmőes a uma concentraçăo de 80%, baixa com o
início da respiraçăo da mistura gasosa do balăo, onde se encontrava numa
concentraçăo de apenas 45%. Seguem-se alguns ciclos ventilatórios em que há
uma progressiva aproximaçăo da concentraçăo do N2 que entra e que
sai dos pulmőes. Poucos segundos depois do início do teste, o traçado
estabiliza, mostrando, a partir daí, uma pequena e constante diferença entre a
concentraçăo de N2 inspirada e expirada. Este ponto indica a
obtençăo de equilíbrio gasoso entre o balăo e os pulmőes. A pequena
diferença remanescente, entre o N2 do ar que entra e do ar que sai
dos pulmőes, deve-se ao fato do ar que sai ter uma temperatura superior ao do
ar que entra. Com a definiçăo deste ponto obtivemos a primeira resposta que
buscávamos: o momento em que a mensuraçăo do débito pode ser iniciada.
Como se vę na figura 5, o nitrogęnio, que imediatamente antes
do teste se encontrava nos pulmőes a uma concentraçăo de 80%, baixa com o
início da respiraçăo da mistura gasosa do balăo, onde se encontrava numa
concentraçăo de apenas 45%. Seguem-se alguns ciclos ventilatórios em que há
uma progressiva aproximaçăo da concentraçăo do N2 que entra e que
sai dos pulmőes. Poucos segundos depois do início do teste, o traçado
estabiliza, mostrando, a partir daí, uma pequena e constante diferença entre a
concentraçăo de N2 inspirada e expirada. Este ponto indica a
obtençăo de equilíbrio gasoso entre o balăo e os pulmőes. A pequena
diferença remanescente, entre o N2 do ar que entra e do ar que sai
dos pulmőes, deve-se ao fato do ar que sai ter uma temperatura superior ao do
ar que entra. Com a definiçăo deste ponto obtivemos a primeira resposta que
buscávamos: o momento em que a mensuraçăo do débito pode ser iniciada.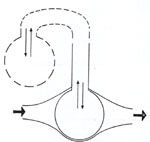 O traçado de N2O mostra, a partir do momento em que
o equilíbrio gasoso é alcançado, um curso francamente descendente, indicando
a absorçăo deste gás pelo sangue que perfunde os pulmőes. Alguns segundos
após o início deste traçado descendente, observa-se uma alteraçăo do plano
por ele seguido. A curva descendente toma-se menos inclinada, ou seja, mais
próxima da horizontal. A mudança de inflexăo desta curva foi por nós
interpretada como indicando o momento em que começa a recirculaçăo de sangue
nos pulmőes. Para comprovar a exatidăo desta interpretaçăo, fizemos
experięncias oferecendo, por broncoespirometria, a mistura gasosa usada no
teste apenas a um pulmăo, enquanto o pulmăo contralateral recebia ar ambiente.
O que observamos pode ser assim resumido (fig. 6): o traçado obtido a partir do
pulmăo que recebera a mistura gasosa usual para o teste, era em tudo semelhante
ao representado na figura 5. O traçado obtido para o pulmăo que recebera ar
ambiente, mostrava composiçăo compatível com ar. Este pulmăo, no entanto,
alguns segundos depois de iniciado o registro, passava a exalar N2O
em concentraçăo crescente. O ponto de aparecimento de N2O neste
pulmăo coincidia com o momento da mudança da inflexăo da curva de N2O
no outro pulmăo. O que confirmava a interpretaçăo de que esta mudança de
inflexăo indicava o início da recirculaçăo sangüínea, ou seja, o momento
até o qual a mensuraçăo do débito era confiável 12.
O traçado de N2O mostra, a partir do momento em que
o equilíbrio gasoso é alcançado, um curso francamente descendente, indicando
a absorçăo deste gás pelo sangue que perfunde os pulmőes. Alguns segundos
após o início deste traçado descendente, observa-se uma alteraçăo do plano
por ele seguido. A curva descendente toma-se menos inclinada, ou seja, mais
próxima da horizontal. A mudança de inflexăo desta curva foi por nós
interpretada como indicando o momento em que começa a recirculaçăo de sangue
nos pulmőes. Para comprovar a exatidăo desta interpretaçăo, fizemos
experięncias oferecendo, por broncoespirometria, a mistura gasosa usada no
teste apenas a um pulmăo, enquanto o pulmăo contralateral recebia ar ambiente.
O que observamos pode ser assim resumido (fig. 6): o traçado obtido a partir do
pulmăo que recebera a mistura gasosa usual para o teste, era em tudo semelhante
ao representado na figura 5. O traçado obtido para o pulmăo que recebera ar
ambiente, mostrava composiçăo compatível com ar. Este pulmăo, no entanto,
alguns segundos depois de iniciado o registro, passava a exalar N2O
em concentraçăo crescente. O ponto de aparecimento de N2O neste
pulmăo coincidia com o momento da mudança da inflexăo da curva de N2O
no outro pulmăo. O que confirmava a interpretaçăo de que esta mudança de
inflexăo indicava o início da recirculaçăo sangüínea, ou seja, o momento
até o qual a mensuraçăo do débito era confiável 12. Na abordagem que agora faço, o ponto que merece destaque, em
relaçăo ŕ técnica recém descrita é a verificaçăo, por ela permitida, de
que o fluxo de sangue pelos pulmőes é influenciado pelos movimentos
ventilatórios. Assim, pude verificar, usando um volume de ar corrente entre
duas a tręs vezes o volume normal e empregando freqüęncias respiratórias
crescentes, que o fluxo de sangue pelos pulmőes aumentava com o aumento da
freqüęncia respiratória. A figura 7 mostra este fenômeno em oito pacientes
nos quais se determinou o fluxo sangüíneo pulmonar, a diferentes freqüęncias
respiratórias. Há uma clara relaçăo entre esta freqüęncia e o fluxo
observado. A 70 movimentos ventilatórios por minuto, o fluxo sangüíneo
pulmonar
Na abordagem que agora faço, o ponto que merece destaque, em
relaçăo ŕ técnica recém descrita é a verificaçăo, por ela permitida, de
que o fluxo de sangue pelos pulmőes é influenciado pelos movimentos
ventilatórios. Assim, pude verificar, usando um volume de ar corrente entre
duas a tręs vezes o volume normal e empregando freqüęncias respiratórias
crescentes, que o fluxo de sangue pelos pulmőes aumentava com o aumento da
freqüęncia respiratória. A figura 7 mostra este fenômeno em oito pacientes
nos quais se determinou o fluxo sangüíneo pulmonar, a diferentes freqüęncias
respiratórias. Há uma clara relaçăo entre esta freqüęncia e o fluxo
observado. A 70 movimentos ventilatórios por minuto, o fluxo sangüíneo
pulmonar 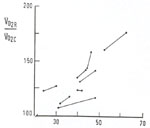 alcança, praticamente, o dobro do valor observado a uma freqüęncia
de repouso. Este aumento é realmente notável considerando-se que o seu grande
propulsor é a bomba ventilatória. Estudos subseqüentes6,7
mostraram que este aumento também se verifica durante exercício, se o
indivíduo aumentar voluntariamente a freqüęncia ventilatória que vinha,
espontaneamente, empregando. A percentagem. de aumento obtida é, no entanto,
menor que em repouso e se torna cada vez menor ŕ medida que o exercício se
torna mais intenso (tab. 1 e fig. 8). Como se observa na tabela I, utilizando-se
da mesma freqüęncia ventilatória forçada (cerca de 30 ciclos ventilatórios
por minuto) e um mesmo volume de ar corrente (cerca de tręs vezes o volume de
ar corrente espontâneo) e superpondo-se este regime ventilatório ŕquele
normalmente observado em repouso e a 400, 800 e 1200 kpm/min, verifica-se que
esta manobra ventilatória forçada determina, em repouso, um aumento
alcança, praticamente, o dobro do valor observado a uma freqüęncia
de repouso. Este aumento é realmente notável considerando-se que o seu grande
propulsor é a bomba ventilatória. Estudos subseqüentes6,7
mostraram que este aumento também se verifica durante exercício, se o
indivíduo aumentar voluntariamente a freqüęncia ventilatória que vinha,
espontaneamente, empregando. A percentagem. de aumento obtida é, no entanto,
menor que em repouso e se torna cada vez menor ŕ medida que o exercício se
torna mais intenso (tab. 1 e fig. 8). Como se observa na tabela I, utilizando-se
da mesma freqüęncia ventilatória forçada (cerca de 30 ciclos ventilatórios
por minuto) e um mesmo volume de ar corrente (cerca de tręs vezes o volume de
ar corrente espontâneo) e superpondo-se este regime ventilatório ŕquele
normalmente observado em repouso e a 400, 800 e 1200 kpm/min, verifica-se que
esta manobra ventilatória forçada determina, em repouso, um aumento  do débito
sangüíneo de 46%, a 400 kpm/min um aumento de 17%, a 800 kpm/min um aumento de
10%, e a 1200 kpm/min um aumento de apenas 4%.
do débito
sangüíneo de 46%, a 400 kpm/min um aumento de 17%, a 800 kpm/min um aumento de
10%, e a 1200 kpm/min um aumento de apenas 4%. A circulaçăo pulmonar, como um todo, é constituída por um
compartimento vascular, ritmicamente modificado em suas dimensőes pelos
movimentos ventilatórios, e limitado, de um lado, pela válvula pulmonar, que
năo permite o retrocesso do sangue e, de outro lado, pela válvula mitral, que
lhe dá fácil vazăo.
A circulaçăo pulmonar, como um todo, é constituída por um
compartimento vascular, ritmicamente modificado em suas dimensőes pelos
movimentos ventilatórios, e limitado, de um lado, pela válvula pulmonar, que
năo permite o retrocesso do sangue e, de outro lado, pela válvula mitral, que
lhe dá fácil vazăo.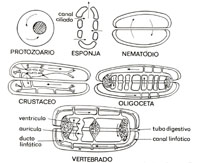 Em seres vivos já com cavidades celômicas ou pseudocelômicas,
como nos nematódios, a circulaçăo é assegurada através da movimentaçăo do
líquido contido nestas cavidades, pela contraçăo periódica de seus músculos
somáticos.
Em seres vivos já com cavidades celômicas ou pseudocelômicas,
como nos nematódios, a circulaçăo é assegurada através da movimentaçăo do
líquido contido nestas cavidades, pela contraçăo periódica de seus músculos
somáticos. Assim, quando os vasos apareceram, nenhuma espécie, acima dos
crustáceos, ficou sem eles. Quando o circuito vascular começou a fechar, com
os oligocetas, o fechamento năo mais retrocedeu; pelo contrário, tornou-se
progressivamente maior. Quando o sistema vagal apareceu, nos peixes mais evoluídos,
năo mais se ausentou. E o mesmo ocorreu com o simpático. E o mesmo com os
linfáticos. E o mesmo com as válvulas venosas.
Assim, quando os vasos apareceram, nenhuma espécie, acima dos
crustáceos, ficou sem eles. Quando o circuito vascular começou a fechar, com
os oligocetas, o fechamento năo mais retrocedeu; pelo contrário, tornou-se
progressivamente maior. Quando o sistema vagal apareceu, nos peixes mais evoluídos,
năo mais se ausentou. E o mesmo ocorreu com o simpático. E o mesmo com os
linfáticos. E o mesmo com as válvulas venosas.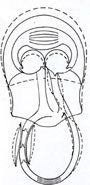 "coraçăo abdominal"
constituído pela veia cava inferior, limitada, de um lado, pelas válvulas
femorais mais altas, e, de outro, pela válvula tricúspide e sujeita a
variaçőes rítmicas da pressăo que a cerca, variaçőes estas oriundas dos
movimentos ventilatórios do diafragma; o "coraçăo ventricular
direito", constituído pelo miocárdio ventricular direito, apoiado pela
aurícula direita, limitado, de um lado, pela válvula tricúspide e, de outro,
pela válvula pulmonar; o "coraçăo pulmonar", constituído pelo
circuito vascular pulmonar, limitado num extremo pela válvula pulmonar e,
noutro, pela válvula mitral, e sujeito ŕs variaçőes rítmicas de volume e de
pressăo impostas pelo fole torácico; o "coraçăo ventricular
esquerdo", constituído pelo miocárdio ventricular esquerdo, apoiado pela
aurícula esquerda, limitado, de um lado, pela válvula mitral e, de outro, pela
válvula aórtica; e, finalmente, o "coraçăo arterial", constituído
pela aorta e seus ramos, limitado, num extremo pela válvula aórtica e, noutro,
pelas primeiras válvulas venosas, acionado pela retraçăo elástica da parede
arterial, ritmicamente distendida pela sístole ventricular.
"coraçăo abdominal"
constituído pela veia cava inferior, limitada, de um lado, pelas válvulas
femorais mais altas, e, de outro, pela válvula tricúspide e sujeita a
variaçőes rítmicas da pressăo que a cerca, variaçőes estas oriundas dos
movimentos ventilatórios do diafragma; o "coraçăo ventricular
direito", constituído pelo miocárdio ventricular direito, apoiado pela
aurícula direita, limitado, de um lado, pela válvula tricúspide e, de outro,
pela válvula pulmonar; o "coraçăo pulmonar", constituído pelo
circuito vascular pulmonar, limitado num extremo pela válvula pulmonar e,
noutro, pela válvula mitral, e sujeito ŕs variaçőes rítmicas de volume e de
pressăo impostas pelo fole torácico; o "coraçăo ventricular
esquerdo", constituído pelo miocárdio ventricular esquerdo, apoiado pela
aurícula esquerda, limitado, de um lado, pela válvula mitral e, de outro, pela
válvula aórtica; e, finalmente, o "coraçăo arterial", constituído
pela aorta e seus ramos, limitado, num extremo pela válvula aórtica e, noutro,
pelas primeiras válvulas venosas, acionado pela retraçăo elástica da parede
arterial, ritmicamente distendida pela sístole ventricular. reservatório de pressăo, potencializador do trabalho
ventricular. A croça da aorta e os seus grandes ramos possuem várias analogias
com este "bulbus arteriosus". Por outro lado, há evidęncia de que a
perda de elasticidade das grandes artérias reduz a eficięncia circulatória,
como um todo.
reservatório de pressăo, potencializador do trabalho
ventricular. A croça da aorta e os seus grandes ramos possuem várias analogias
com este "bulbus arteriosus". Por outro lado, há evidęncia de que a
perda de elasticidade das grandes artérias reduz a eficięncia circulatória,
como um todo.